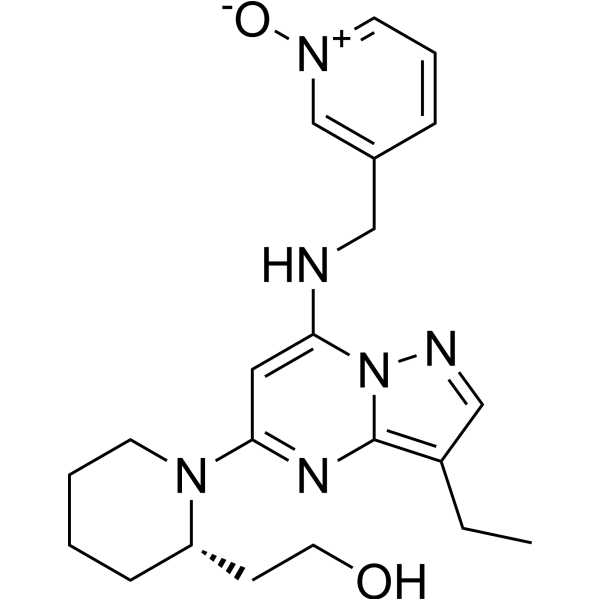
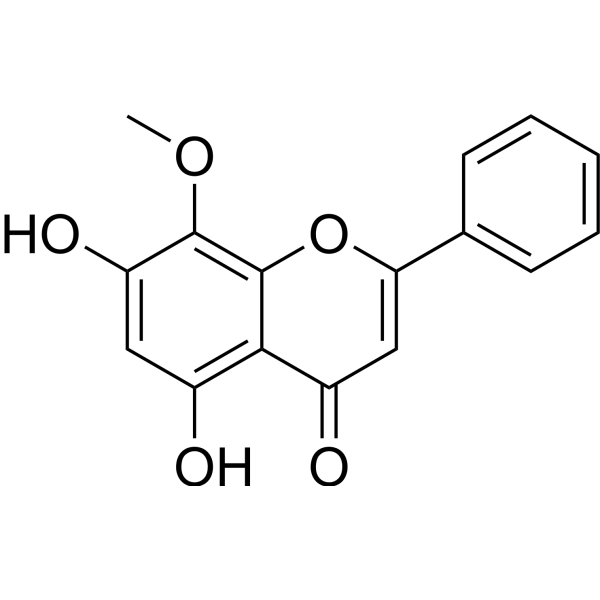
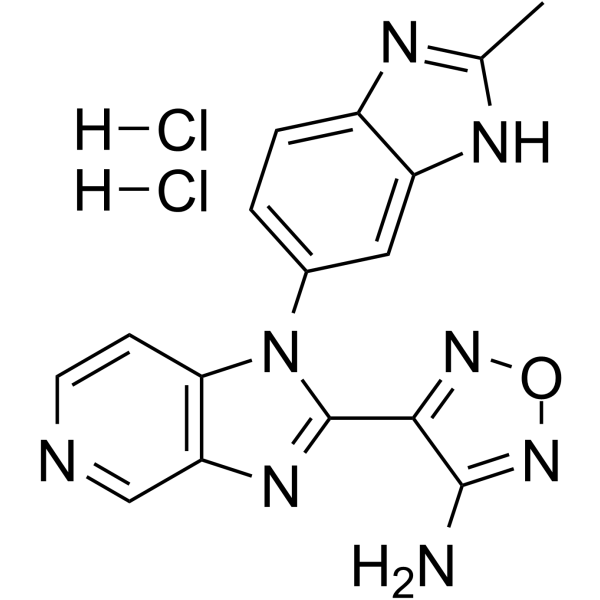
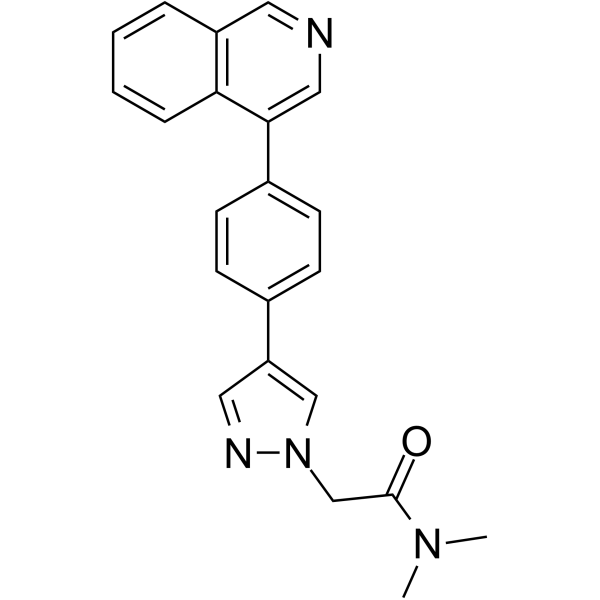
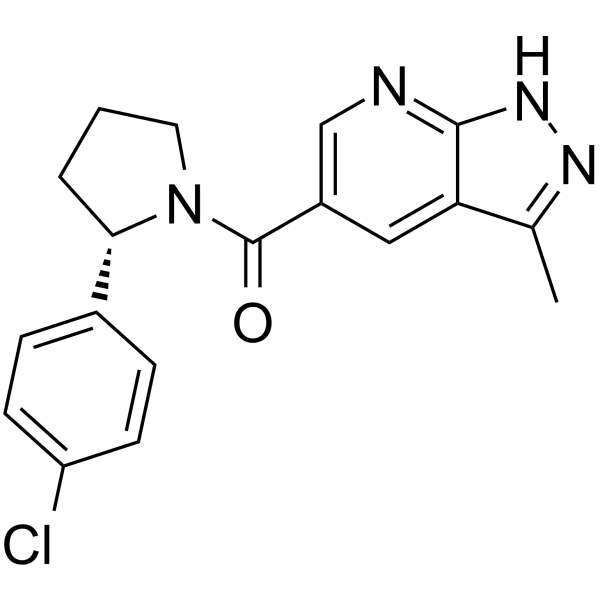
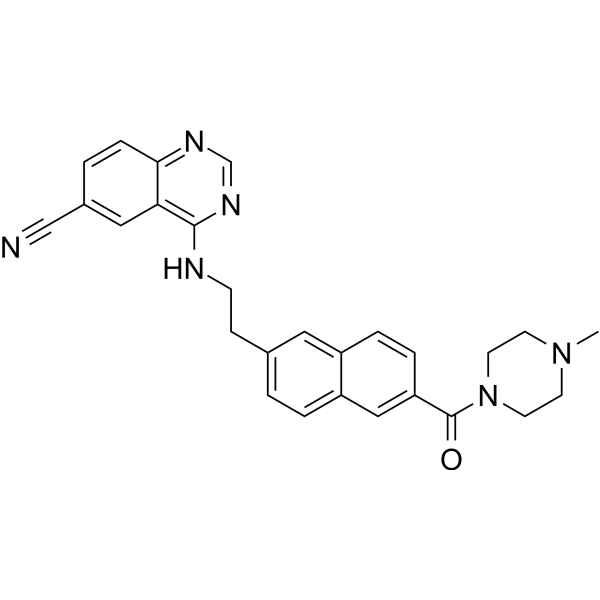
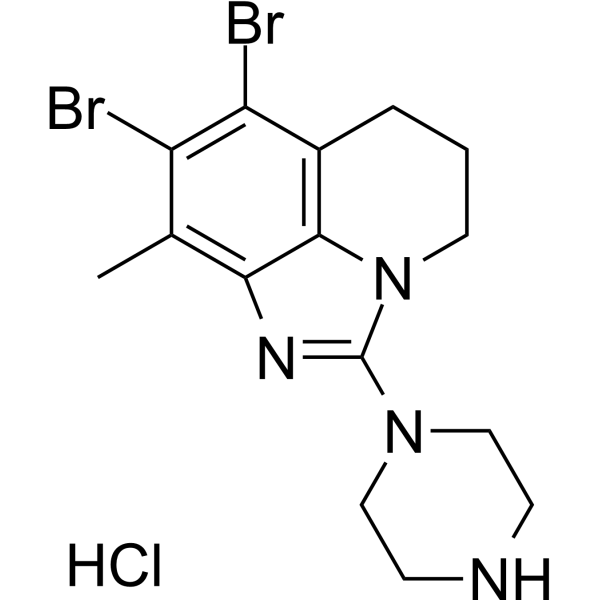
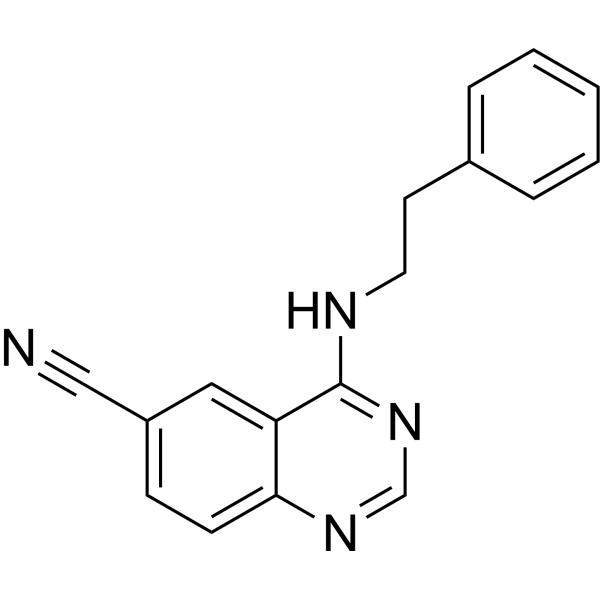
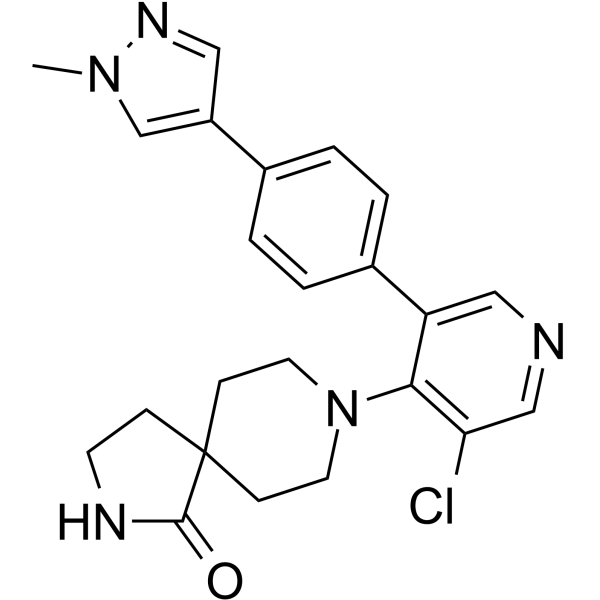
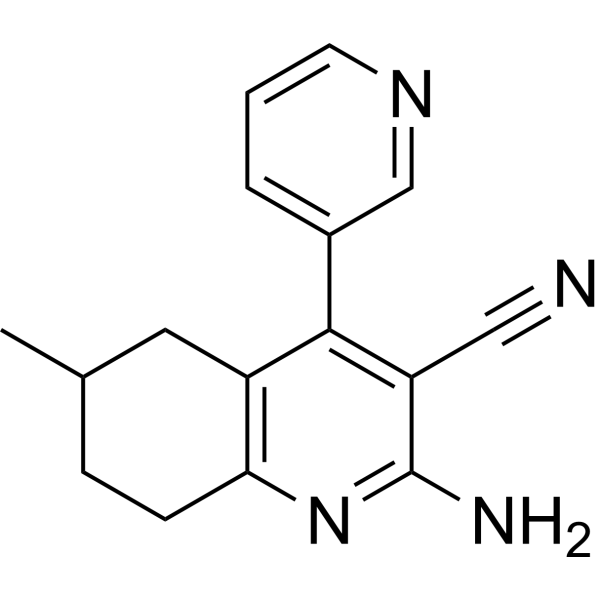
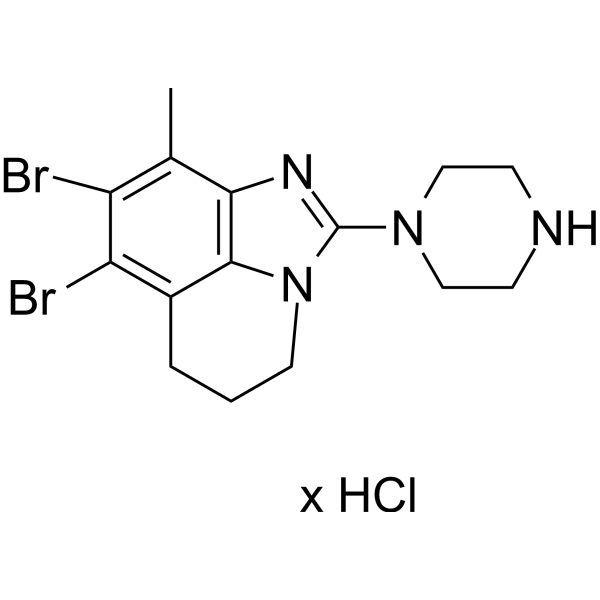
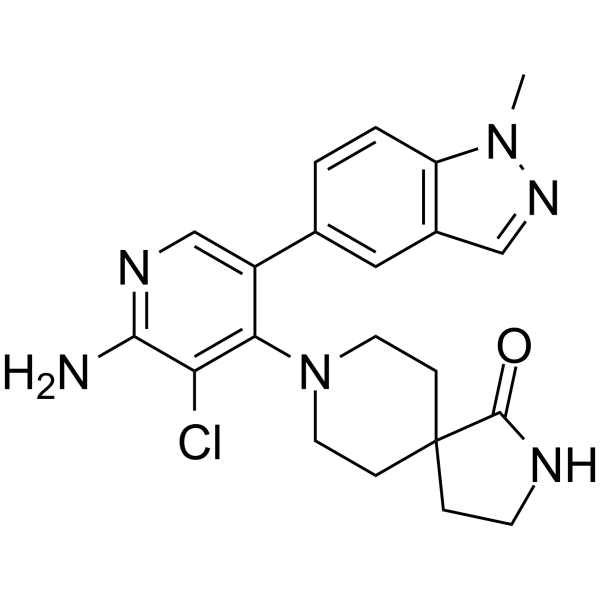
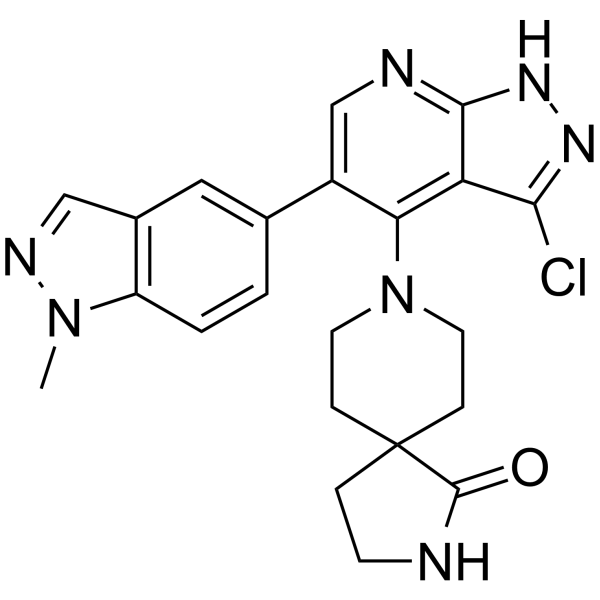
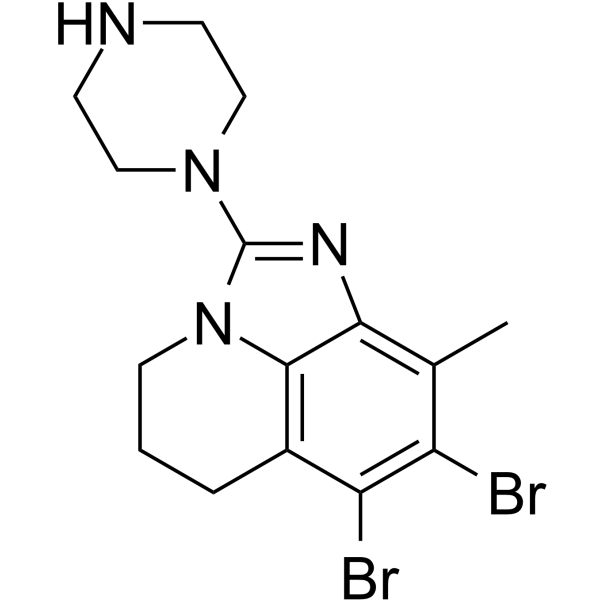
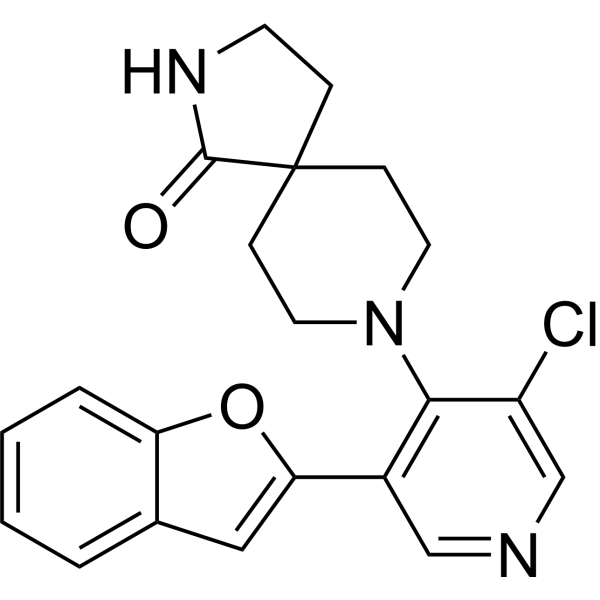
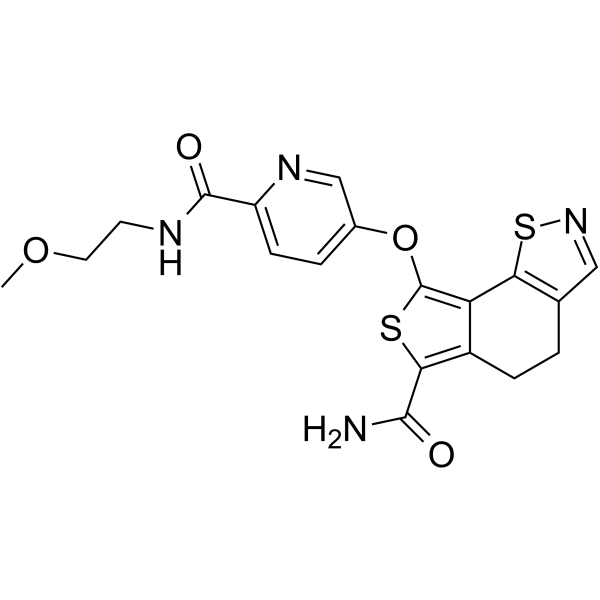
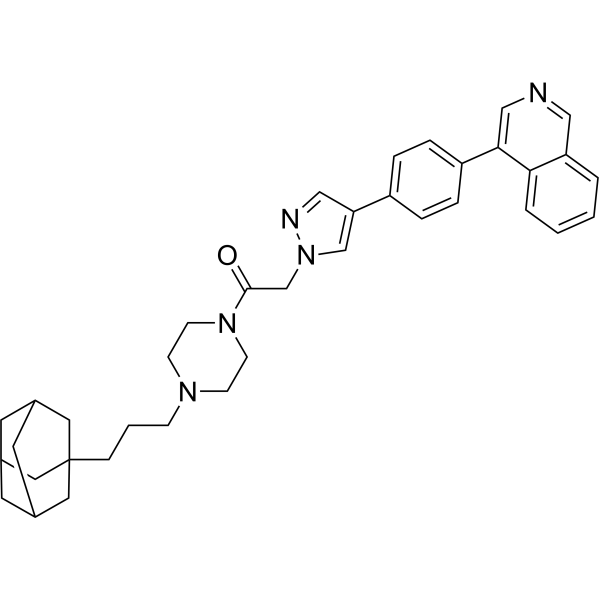
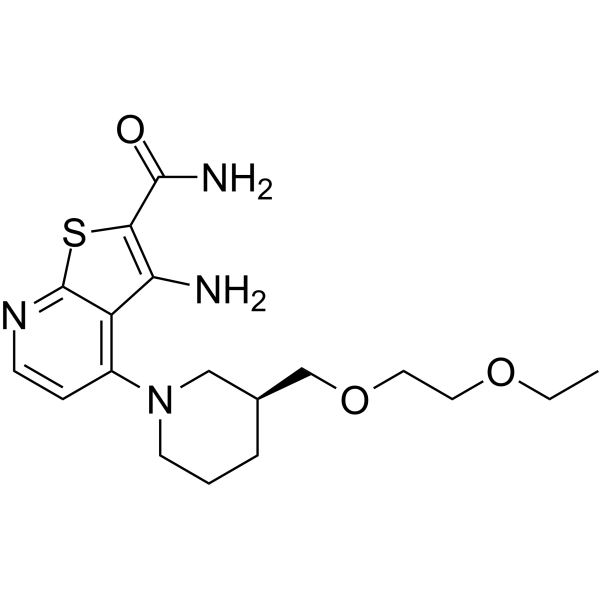
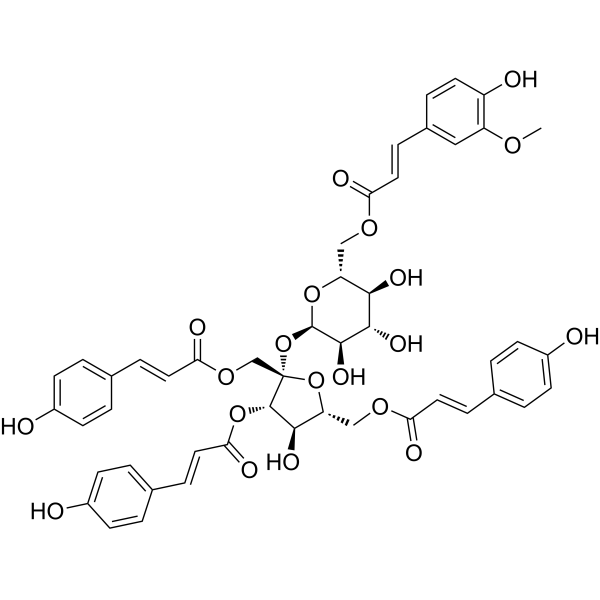
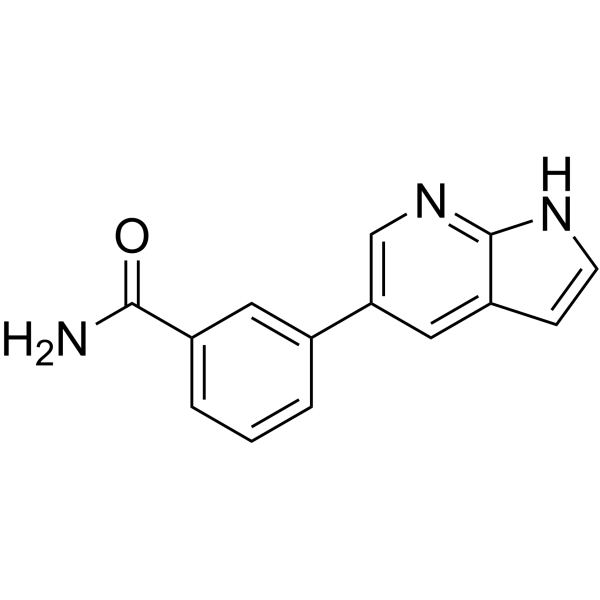
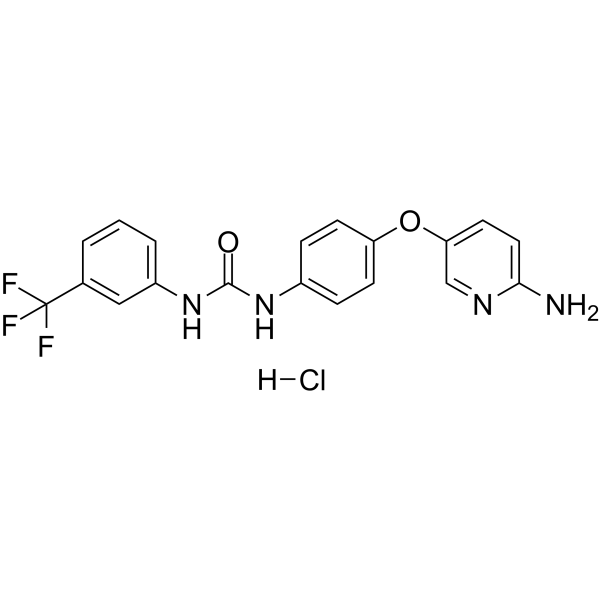
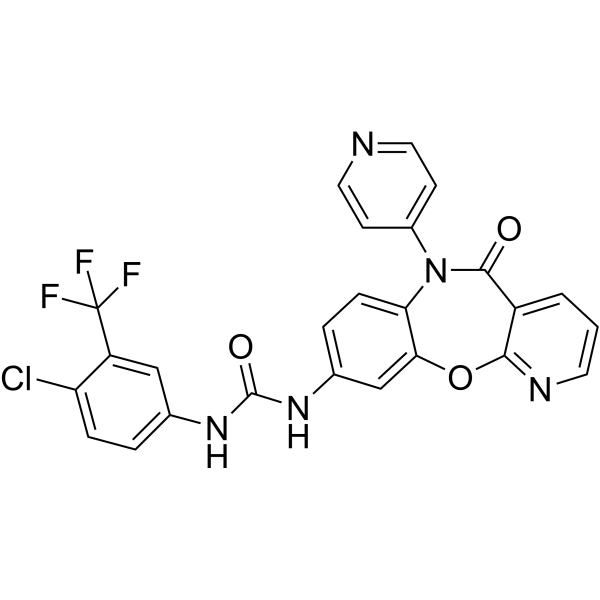
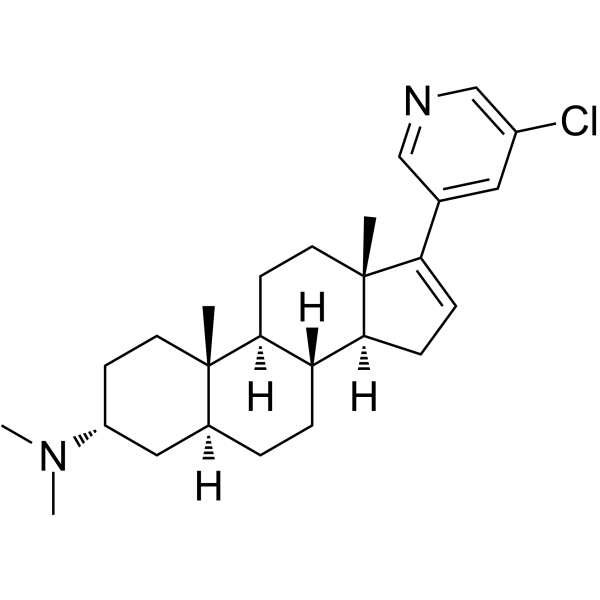
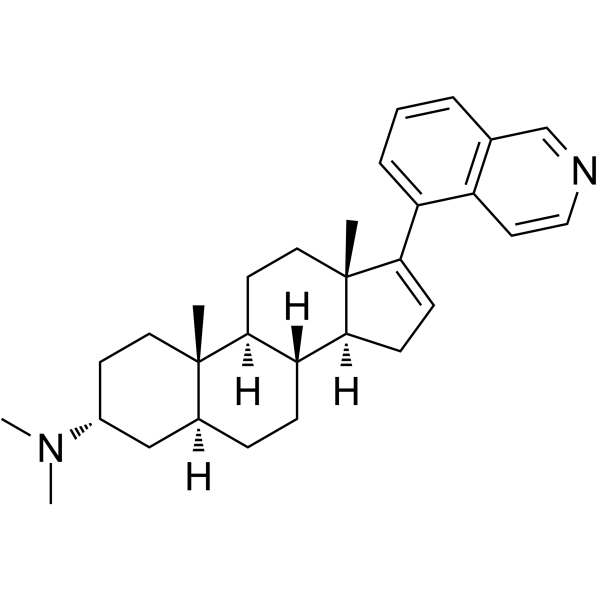
From 11:00 pm to 12:00 pm EST ( 8:00 pm to 9:00 pm PST ) on January 6th, the website will be under maintenance. We are sorry for the inconvenience. Please arrange your schedule properly.
AS2863619 enables conversion of antigen-specific effector/memory T cells into Foxp3 + regulatory T (Treg) cells for the treatment of various immunological diseases. AS2863619 is a potent, orally active cyclin-dependent kinase 8 (CDK8) and CDK19 inhibitor with IC50s of 0.61 nM and 4.28 nM, respectively. STAT5 activation enhanced by AS2863619 inhibition of CDK8/19, which consequently activates the Foxp3 gene .
SEL120-34A monohydrochloride is an ATP-competitive and selective CDK8 inhibitor, inhibits kinase activities of CDK8/CycC and CDK19/CycC complexes with IC50s of 4.4 nM and 10.4 nM, respectively, with a Kd of 3 nM for CDK8. SEL120-34A monohydrochloride weakly inhibits CDK9 (calculated IC50=1070 nM), but shows no obvious activity against CDK1, 2, 4, 6, 5, 7. SEL120-34A monohydrochloride inhibits phosphorylation of STAT1 S727 and STAT5 S726 . Has anti-tumor activity .
Senexin A is an inhibitor of CDK8/19 (IC50: 280 nM, CDK8) and an inhibitor downstream of p21 transcription. It only inhibits p21-induced transcription but does not inhibit other biological effects of p21. Senexin A inhibits CMV-GFP induction as well as the p21 stimulatory activity of the consensus NF-κB-dependent promoters .
CCT251545 is an orally bioavailable and potent inhibitor of WNT signaling with an IC50 of 5 nM in 7dF3 cells . CCT251545 is a selective chemical probe for exploring the role of CDK8 and CDK19 in human disease .
BRD6989, an analog of the natural product cortistatin A (dCA), inhibits CDK8 and upregulates IL-10. BRD6989 selectively binds a complex of CDK8 with an IC50 of ~200 nM. BRD6989 inhibits the kinase activity of recombinant CDK8 or CDK19 complexes .
SEL120-34A hydrochloride is a potent, selective, orally available, ATP-competitive CDK8 inhibitor, with IC50s of 4.4 nM and 10.4 nM for CDK8/CycC and CDK19/CycC, respectively, with antitumor activity.
JH-XVI-178 is a highly efficient and selective CDK8/19 inhibitor, with IC50 values of 1 and 2 nM for CDK8 and CDK19, respectively. JH-XVI-178 has a low clearance rate and moderate oral pharmacokinetic properties .
SEL120-34A is a potent, selective, orally available, ATP-competitive CDK8 inhibitor, with IC50s of 4.4 nM and 10.4 nM for CDK8/CycC and CDK19/CycC, respectively, with antitumor activity.
CDK8-IN-12 is an orally active, potent CDK8 inhibitor with a Ki of 14 nM. CDK8-IN-12 has off-target kinase inhibition on GSK-3α, GSK-3β, PCK-θ with Kis of 13 nM, 4 nM, 109 nM, respectively. CDK8-IN-12 shows potent anti-proliferative effects selectively on MV4-11 cell. CDK8-IN-12 is an anti-cancer agent .
CDK8/19-IN-1 is a potent, selective and oral bioavailable CDK8/19 dual inhibitor, with IC50s of 0.46 nM, 0.99 nM and 270 nM for CDK8, CDK19 and CDK9, respectively.
LY2857785 is a type I reversible and competitive ATP kinase inhibitor against CDK9 (IC50 11 nM) and other transcription kinases CDK8 (IC50 16 nM), and CDK7 (IC50 246 nM).
3MB-PP1, a bulky purine analog, is a Polo-like kinase 1 (Plk1) inhibitor. 3MB-PP1 blocks mitotic progression and cell division arise through target Plk1 in in cells expressing analog-sensitive Plk1 alleles. 3MB-PP1 specifically inhibits the activity of analog-sensitive Ssn3(Cdk8). 3MB-PP1 inhibits Leu93 Mutant Zipper-interacting protein kinase (Leu93-ZIPK; IC50=2 μM). 3MB-PP1 can be used for the research of hypha formation of Candida albicans and cell division .
CDK8-IN-11 is a potent and selective CDK8 inhibitor with an IC50 value of 46 nM. CDK8-IN-11 inhibits WNT/β-catenin signaling pathway. CDK8-IN-11 can be used in the research of colon cancer .
JH-XI-10-02 is a PROTAC connected by ligands for Cereblon and CDK. JH-XI-10-02 is a highly potent and selective PROTAC CDK8 degrader, with an IC50 of 159 nM. JH-XI-10-02 causes proteasomal degradation, does not affect CDK8 mRNA levels. JH-XI-10-02 shows no effect on CDK19 .
AS2863619 free base enables conversion of antigen-specific effector/memory T cells into Foxp3 + regulatory T (Treg) cells for the treatment of various immunological diseases. AS2863619 free base is a potent, orally active cyclin-dependent kinase 8 (CDK8) and CDK19 inhibitor with IC50s of 0.61 nM and 4.28 nM, respectively. STAT5 activation enhanced by AS2863619 free base inhibition of CDK8/19, which consequently activates the Foxp3 gene .
Senexin C is a selective and orally active CDK8/19 inhibitor. Senexin C shows a strong tumor-enrichment pharmacokinetic (PK) profile and tumor-pharmacodynamic (PD) marker responses. Senexin C inhibits the growth of MV4-11 leukemia cells with good tolerability .
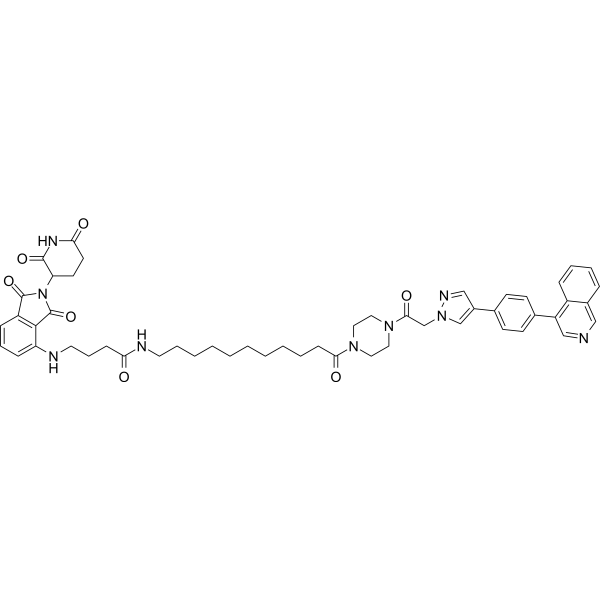
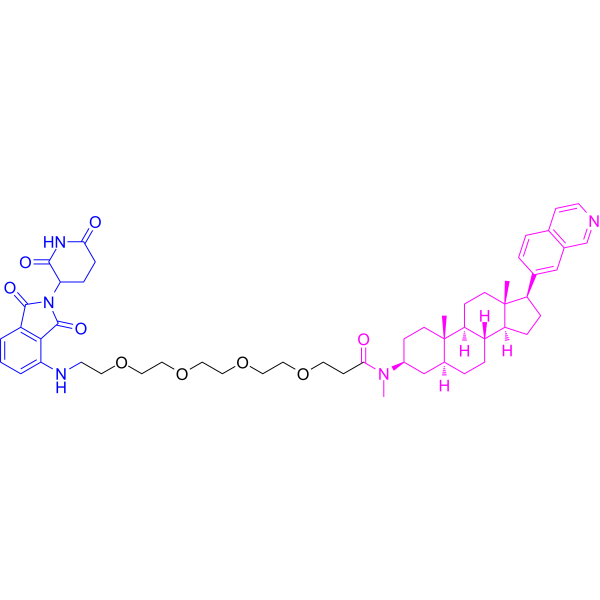
LL-K8-22 is a potent, selective and durable PROTAC CDK8-cyclin C dual degrader, with DC50 values of 2.52 and 2.64 μM, respectively. LL-K8-22 also suppresses STAT1 Ser 727 phosphorylation. LL-K8-22 inhibits E2F- and MYC-driven carcinogenic transcriptional programs. LL-K8-22 can be used for triplenegative breast cancer (TNBC) research. (Pink: Ligand for target protein (HY-168683); Black: Linker (HY-Y0340); Blue: Ligand for E3 ligase (HY-N2427)) .
Vanicoside B is a phenylpropanoyl sucrose derivative, can be isolated from the herb Persicaria dissitiflora. Vanicoside B targets cyclin-dependent kinase 8(CDK8) and exhibits anti-tumor activity. The potential mechanism is Vanicoside B blocks CDK8-mediated signaling pathways and decreases the expression of epithelial-mesenchymal transition proteins, so that it leads to cell cycle arrest and apoptosis .
CDK8-IN-13 is a potent, selective and orally active CDK8 inhibitor with an IC50 value of 51.9 nM. CDK8-IN-13 induces apoptosis. CDK8-IN-13 decreases the expression of p-STAT1 S727 and p-STAT5 S726. CDK8-IN-13 shows antitumor activity .
MSC2530818 (Standard) is the analytical standard of MSC2530818. This product is intended for research and analytical applications. MSC2530818 is a potent, selective and orally available CDK8 inhibitor with an IC50 of 2.6 nM for CDK8.
CDK8 Human Pre-designed siRNA Set A contains three designed siRNAs for CDK8 gene (Human), as well as a negative control, a positive control, and a FAM-labeled negative control.
CDK8-IN-5 is a potent CDK8 inhibitor with an IC50 of 72 nM. CDK8-IN-5 shows anti-inflammatory activities with 43% IL-10 enhancement rate. CDK8-IN-5 has the potential for the research of inflammatory bowel disease .
CDK8/19-IN-2 (compound 12) is an orally active and potent cyclin-dependent kinase 8/19 (CDK8 and CDK19) inhibitor, with IC50 values of 2.08 and 2.49 nM, respectively. CDK8/19-IN-2 can be used for acute myeloid leukemia (AML), breast cancer, and lymphoma research .
CDK8-IN-19 (Compound 3d) is a CDK8 inhibitor with an IC50 of 25.08 nM. CDK8-IN-19 has a broad-spectrum anticancer activity, such as leukemia, melanoma and breast cancer, without significant cytotoxic effect on the normal Vero cells (mean IC50s of 2.87, 3.65, 3.79 and 45.48 μM, respectively) .
Cdk8 Rat Pre-designed siRNA Set A contains three designed siRNAs for Cdk8 gene (Rat), as well as a negative control, a positive control, and a FAM-labeled negative control.
Cdk8 Mouse Pre-designed siRNA Set A contains three designed siRNAs for Cdk8 gene (Mouse), as well as a negative control, a positive control, and a FAM-labeled negative control.
JH-VIII-49 (compound 10) is a potent and selective CDK8 inhibitor (IC50=16 nM) with excellent biological activity. JH-VIII-49 promotes CDK8 inhibition through its steroid backbone design. JH-VIII-49 can be used in the synthesis of PROTAC as a target protein ligand of JH-XI-10-02 (HY-111518) .
CDK8-IN-16 (Compound 51) is an orally active dual inhibitor for CDK8 and CDK19 wih IC50 of 5.1 nM and 5.6 nM. CDK8-IN-16 inhibits the phospho-STAT1 SER727 with an IC50 of 17.9 nM in SW620 cell, inhibits WNT signaling pathway with an IC50 of 7.2 nM in 7dF3 cell. CDK8-IN-16 exhibits good pharmacokinetic characteristics in rat models with an oral bioavailability of 57% .
CDK8-IN-9 (compound 22) is a potent type II CDK8 inhibitor with an IC50 value of 48.6 nM. CDK8-IN-9 can inhibit tumor growth and is used in colorectal cancer studies .
CDK8-IN-11 hydrochloride is a potent and selective CDK8 inhibitor with an IC50 value of 46 nM. CDK8-IN-11 hydrochloride inhibits WNT/β-catenin signaling pathway. CDK8-IN-11 hydrochloride can be used in the research of colon cancer .
CDK8-IN-10 (compound 2) is a potent, selective cyclin-dependent kinase (CDK8) inhibitor with an IC50 value of 8.25 nM. CDK8-IN-10 can be used for research of cancer .
P162-0948 is a selective CDK8 inhibitor with an IC50 value of 50.4 nM. P162-0948 reduces cell migration and protein expression of EMT-related proteins in A549 human alveolar epithelial cell lines. P162-0948 reduces phosphorylation of Smad, which suggests disruption of the TGF-β/Smad signaling pathway. P162-0948 is promising for research of pulmonary fibrosis .
CDK8/19-IN-3 (compound 3-7) is a potent and selective CDK8 and CDK19 inhibitor. CDK8/19-IN-3 upregulates IL-10 levels. CDK8/19-IN-3 has the potential for the research of inflammatory bowel disease (IBD) .
CDK8-IN-6 (compound 9) is a potent cyclin-dependent kinase 8(CDK8) inhibitor with an Kd of 13 nM. CDK8-IN-6 shows cytotoxicity for MOLM-13, OCI-AML3, MV4-11, NRK and H9c2 cells with IC50s of 11.2, 7.5, 8.6, 20.5, 12.5-25 µM, respectively. CDK8-IN-6 has the potential for the research of AML-cancer .
CDK8-IN-7 (compound 12) is a potent and selective cyclin-dependent kinase 8(CDK8) inhibitor with an Kd of 3.5 nM. CDK8-IN-7 shows cytotoxicity for MOLM-13, OCI-AML3, MV4-11, NRK and H9c2 cells with IC50s of 5.9, 4.8, 5.4, 16.2, 12.5-25 µM, respectively. CDK8-IN-7 has the potential for the research of AML-cancer .
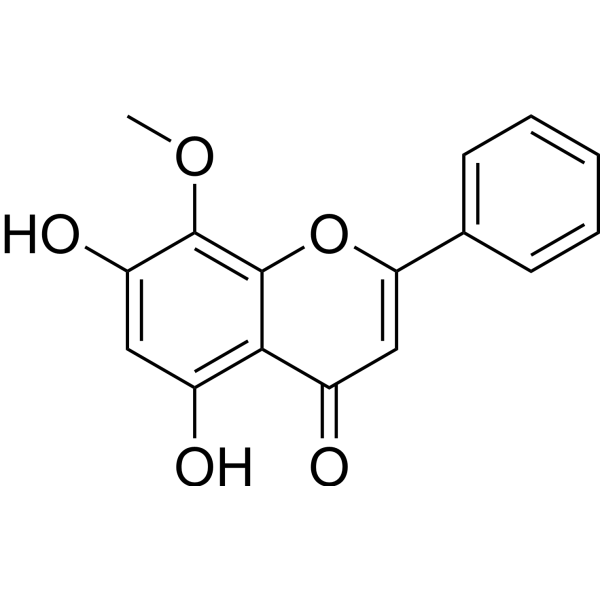
Wogonin (Standard) is the analytical standard of Wogonin. This product is intended for research and analytical applications. Wogonin is a naturally occurring mono-flavonoid, can inhibit the activity of CDK8 and Wnt, and exhibits anti-inflammatory and anti-tumor effects.
CDK8-IN-15 (Compound 46) is a potent CDK8 inhibitors with an IC50 value of 57 nM. It can enhance the thermal stability of CDK8 along with inhibition against NF-κB and have favourable selectivity across the CDK family and tyrosine kinase. Additionally, it also demostrates a positive effect in vitro psoriasis model induced by TNF-α and alleviats the inflammatory response enhancing the expression of Foxp3 and IL-10, which is promising for research of psoriasis diseases .
CDK-IN-13 (compound 32E) is a potent and selective inhibitor of CDK12/cyclinK with an IC50 of 3 nM. CDK-IN-13 inhibits the growth of the HER2-positive breast cancer cell lines .
CDK8 modulates the transcriptional output from distinct transcription factors involved in oncogenic control, including the Wnt/β-catenin pathway, Notch, p53, and transforming growth factor β. Abnormal activity ofCDK8 along with its partner protein cyclin C (CycC) is a common feature of many diseases including colorectal cancer. CDK8/CycC Recombinant Human Active Protein Kinase can be used to study the function of CDK8/CycC .
CDK8-IN-14 (compound 12) inhibits CDK8 with an IC50 value of 39.2 nM and has anti-AML cell proliferation activity (molm-13 GC50 = 0.02±0.01μM, MV4-11 GC50 = 0.03±0.01μM) .
Senexin A hydrochloride is an inhibitor of CDK8/19 (IC50: 280 nM, CDK8) and an inhibitor downstream of p21 transcription. It only inhibits p21-induced transcription but does not inhibit other biological effects of p21. Senexin A hydrochloride inhibits CMV-GFP induction as well as the p21 stimulatory activity of the consensus NF-κB-dependent promoters .
CDK6/9-IN-2 is a highly active dual inhibitor of CDK6 (IC50 = 15 nM) and CDK9 (IC50 = 22 nM). CDK6/9-IN-2 is selective for CDK2, CDK8, and CDK11. CDK6/9-IN-2 inhibits the proliferation of HaCaT cells induced by IFN-γ/TNF-α and suppresses the STAT3 pathway and the expression of inflammatory factors. CDK6/9-IN-2 can alleviate psoriatic dermatitis and is useful in psoriasis research .
CDK8-IN-2 is an orally active CDK8 inhibitor of an IC50 values of 0.010 μM. CDK8-IN-2 shows a CDK19IC50 value of 0.026 μM. CDK8-IN-2 inhibits phospho-STAT1, a pharmacodynamic biomarker of CDK8. CDK8-IN-2 inhibits WNT pathway activity. CDK8-IN-2 can be used for the research of colorectal carcinoma .
Adamantane-Butyl alcohol is a selective and persistent degrader targeting the CDK8-cyclin C complex. Adamantane-Butyl alcohol is the linker for LL-K8-22 (PROTACCDK8-cyclin C, HY-149209) and HyT .
CDK8-IN-1 (Standard) is the analytical standard of CDK8-IN-1 (HY-103492). This product is intended for research and analytical applications. CDK8-IN-1 is a potent and selective CDK8 inhibitor with an IC50 of 3 nM.
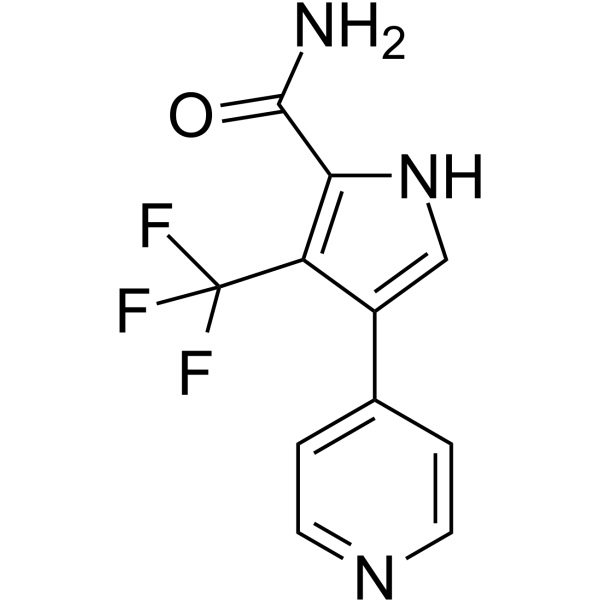
CDK8-IN-20 (Compound 67j) is a selective, potent and orally active type I CDK8 inhibitor with an IC50 of 70.5 nM. CDK8-IN-20 shows IC50 values of 147.8, 726.9 and 217.4 nM for homologous kinase CDK19, CDK7 and CDK9. CDK8-IN-20 can inhibit the Wnt/β-catenin pathway and downregulate the expression of β-catenin, Cyclin D1, and c-Myc. CDK8-IN-20 can induce ROS production and cause G2/M and S phase arrest. CDK8-IN-20can be used for the research of cancer, such as colon cancer .
RO8323 is an orally active, selective CDK8/CDK19 inhibitor, with an IC50 of 2 nM against CDK8 and 3 nM against CDK19. RO8323 promotes regulatory T cell differentiation, inhibits effector T cell generation, reverses the Teff/Treg ratio, upregulates IL-10 production in myeloid cells, and suppresses the production of TNF-α, IL-6 and IL-12. RO8323 enhances immune reconstitution and prolongs cardiac allograft survival in a dose-dependent manner. RO8323 can be used in the research of chronic graft-versus-host disease, cardiac allograft rejection, acute graft-versus-host disease and experimental autoimmune encephalomyelitis .
Senexin B (Standard) (SNX2-1-165 (Standard)) is the analytical standard of Senexin B (HY-101800). This product is intended for research and analytical applications. Senexin B (SNX2-1-165; BCD-115) is a potent, highly water-soluble and bioavailable CDK8/19 inhibitor, with Kds of 140 nM for CDK8 and 80 nM for CDK19.
Dinaciclib (Standard) is the analytical standard of Dinaciclib (HY-10492). This product is intended for research and analytical applications. Dinaciclib (SCH 727965) is a potent inhibitor of CDK, with IC50s of 1 nM, 1 nM, 3 nM, and 4 nM for CDK2, CDK5, CDK1, and CDK9, respectively .
VCC972839:01 is an inhibitor for cyclin dependent kinase(CDK9), with IC50 of 7 nM. VCC972839:01 inhibits the cell viability of SCLC cells in nanomolar levels. VCC972839:01 induces apoptosis through an intrinsic pathway. VCC972839:01 exhibits antitumor activity in mouse model .
3MB-PP1 (Standard) is the analytical standard of 3MB-PP1 (HY-102069). This product is intended for research and analytical applications. 3MB-PP1, a bulky purine analog, is a Polo-like kinase 1 (Plk1) inhibitor. 3MB-PP1 blocks mitotic progression and cell division arise through target Plk1 in in cells expressing analog-sensitive Plk1 alleles. 3MB-PP1 specifically inhibits the activity of analog-sensitive Ssn3 (Cdk8). 3MB-PP1 inhibits Leu93 Mutant Zipper-interacting protein kinase (Leu93-ZIPK; IC50=2 μM). 3MB-PP1 can be used for the research of hypha formation of Candida albicans and cell division .
Vanicoside B is a phenylpropanoyl sucrose derivative, can be isolated from the herb Persicaria dissitiflora. Vanicoside B targets cyclin-dependent kinase 8(CDK8) and exhibits anti-tumor activity. The potential mechanism is Vanicoside B blocks CDK8-mediated signaling pathways and decreases the expression of epithelial-mesenchymal transition proteins, so that it leads to cell cycle arrest and apoptosis .
Wogonin (Standard) is the analytical standard of Wogonin. This product is intended for research and analytical applications. Wogonin is a naturally occurring mono-flavonoid, can inhibit the activity of CDK8 and Wnt, and exhibits anti-inflammatory and anti-tumor effects.
The CDK8-CCNC-MED12 protein is an important component of the mediator complex, acting as a bridge for gene transcription regulation, transmitting information from gene-specific regulatory factors to RNA polymerase II. It is recruited to the promoter, assembles a preinitiation complex, and promotes transcription. CDK8 Protein, Human ( Unactive, sf9, GST) is the recombinant human-derived CDK8, expressed by Sf9 insect cells, with GST labeled tag. The total length of CDK8 Protein, Human ( Unactive, sf9, GST) is 464 a.a..
The CDK8-CCNC-MED12 protein is an important component of the mediator complex, acting as a bridge for gene transcription regulation, transmitting information from gene-specific regulatory factors to RNA polymerase II. It is recruited to the promoter, assembles a preinitiation complex, and promotes transcription. CDK8 Protein, Human (Sf9) is the recombinant human-derived CDK8, expressed by Sf9 insect cells, with tag-free. The total length of CDK8 Protein, Human (Sf9) is 464 a.a..
The CDK8-CCNC-MED12 protein is an important component of the mediator complex, acting as a bridge for gene transcription regulation, transmitting information from gene-specific regulatory factors to RNA polymerase II. It is recruited to the promoter, assembles a preinitiation complex, and promotes transcription. CDK8-CCNC Protein, Human (Active, sf9, GST) is the recombinant human-derived CDK8-CCNK, expressed by Sf9 insect cells, with N-GST labeled tag.
The CDK8-CCNC-MED12 protein is an important component of the mediator complex, acting as a bridge for gene transcription regulation, transmitting information from gene-specific regulatory factors to RNA polymerase II. It is recruited to the promoter, assembles a preinitiation complex, and promotes transcription. CDK8-CCNC-MED12 Protein, Human (sf9, GST, FLAG, His) is the recombinant human-derived CDK8-CCNC-MED12 protein, expressed by sf9 insect cells , with N-6*His, N-Flag, N-GST labeled tag.
CDK19 plays a key role in cellular homeostasis and developmental programming. CDK19 can interact with p53 to inhibit p53-mediated transcription of p21, and regulates the proliferation of hematopoietic stem cells and acute myeloid leukemia cells. Besides, CDK19 is the paralog of CDK8. CDK8 and CDK19 can cooperate with each other in stimulating NFκB-induced transcription and Dengue virus replication. CDK19-CCNC-MED12 Protein, Human (sf9, GST, Flag, His) is the recombinant human-derived CDK19-CCNC-MED12, expressed by Sf9 insect cells, with N-6*His, N-GST, N-Flag labeled tag.
CDK8 Human Pre-designed siRNA Set A contains three designed siRNAs for CDK8 gene (Human), as well as a negative control, a positive control, and a FAM-labeled negative control.
Cdk8 Rat Pre-designed siRNA Set A contains three designed siRNAs for Cdk8 gene (Rat), as well as a negative control, a positive control, and a FAM-labeled negative control.
Cdk8 Mouse Pre-designed siRNA Set A contains three designed siRNAs for Cdk8 gene (Mouse), as well as a negative control, a positive control, and a FAM-labeled negative control.
Inquiry Online
Your information is safe with us. * Required Fields.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
MedchemExpress Validation 03
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
MedchemExpress Validation 04
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedChemExpress values your privacy and your trust is important to us. We use cookies to enhance your website experience. Some cookies are necessary to run the website.
Privacy and Cookie Policy