From 11:00 pm to 12:00 pm EST ( 8:00 pm to 9:00 pm PST ) on January 6th, the website will be under maintenance. We are sorry for the inconvenience. Please arrange your schedule properly.
hsa-miR-7-5p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
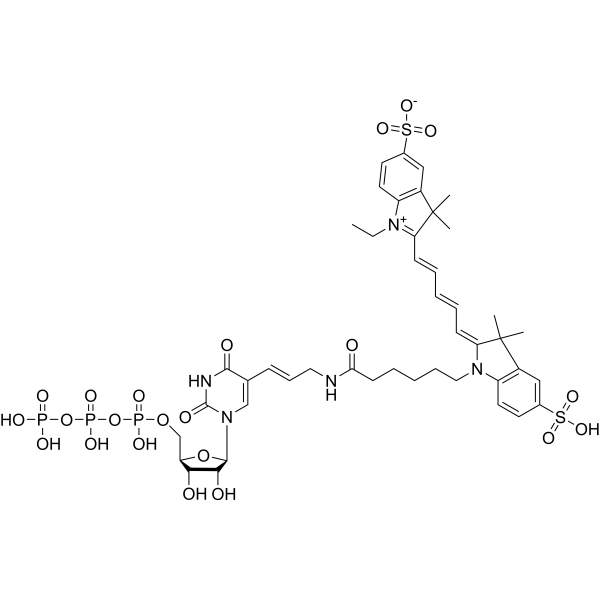
Cy5-UTP is a fluorescently labeled ribonucleotide triphosphate that can be used as a substrate for terminal deoxynucleotidyl transferase (TdT). Cy5-UTP can be employed to label RNA probes generated in vitro (Ex/Em: 650/665 nm). Cy5-UTP can be applied in FISH, multi-color fluorescence analysis, especially in dual-color expression arrays that combine with Cy5-UTP .
hsa-miR-7-5p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-2-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7a-5p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
hsa-miR-7-1-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7b-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7a-2-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7b-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
mmu-miR-7a-2-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-5p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
rno-miR-7b antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-5p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-5p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-5p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
LILRB1 Human Pre-designed siRNA Set A contains three designed siRNAs for LILRB1 gene (Human), as well as a negative control, a positive control, and a FAM-labeled negative control.
rno-miR-7b mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
rno-miR-7b inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
rno-miR-7a-1-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
rno-miR-7b agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-5p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-2-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-1-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
rno-miR-7a-1-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-1-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-2-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7b-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-2-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
rno-miR-7a-1-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-1-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7b-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-2-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-2-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
rno-miR-7a-1-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
Cy5-UTP is a fluorescently labeled ribonucleotide triphosphate that can be used as a substrate for terminal deoxynucleotidyl transferase (TdT). Cy5-UTP can be employed to label RNA probes generated in vitro (Ex/Em: 650/665 nm). Cy5-UTP can be applied in FISH, multi-color fluorescence analysis, especially in dual-color expression arrays that combine with Cy5-UTP .
LILRB1/CD85j/ILT2 Protein is an inhibitory receptor broadly expressed on leukocytes. LILRB1 recognises a wide range of classical HLA-class I allelic variants, as well as the non-classical molecules HLA-F and -G by binding to the conserved a3 domain. LILRB1 also recognises the human CMV-encoded MHC class I homologue UL18. LILRB1 is encoded within the leukocyte receptor complex on 19q13.4. LILRB1 can function as a negative regulator of BiTE molecule-induced tumor cell killing. LILRB1 acts as a novel checkpoint inhibitory molecule capable of restricting BiTE molecule-mediated CD8+ T cell effector function. LILRB1/CD85j/ILT2 Protein, Human (Biotinylated, HEK293, His-Avi) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-Avi, C-6*His labeled tag.
LILRB1/CD85j/ILT2 Protein is an inhibitory receptor broadly expressed on leukocytes. LILRB1 recognises a wide range of classical HLA-class I allelic variants, as well as the non-classical molecules HLA-F and -G by binding to the conserved a3 domain. LILRB1 also recognises the human CMV-encoded MHC class I homologue UL18. LILRB1 is encoded within the leukocyte receptor complex on 19q13.4. LILRB1 can function as a negative regulator of BiTE molecule-induced tumor cell killing. LILRB1 acts as a novel checkpoint inhibitory molecule capable of restricting BiTE molecule-mediated CD8+ T cell effector function. LILRB1/CD85j/ILT2 Protein, Human (HEK293, Fc) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-hFc labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Human (HEK293, His) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-6*His labeled tag.
LILRB1/CD85j/ILT2 Protein is an inhibitory receptor broadly expressed on leukocytes. LILRB1 recognises a wide range of classical HLA-class I allelic variants, as well as the non-classical molecules HLA-F and -G by binding to the conserved a3 domain. LILRB1 also recognises the human CMV-encoded MHC class I homologue UL18. LILRB1 is encoded within the leukocyte receptor complex on 19q13.4. LILRB1 can function as a negative regulator of BiTE molecule-induced tumor cell killing. LILRB1 acts as a novel checkpoint inhibitory molecule capable of restricting BiTE molecule-mediated CD8+ T cell effector function. LILRB1/CD85j/ILT2 Protein, Human (Biotinylated, HEK293, Fc-Avi) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-Avi, C-hFc labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Human (HEK293, mFc) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-mFc labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Human (HEK293, His-Avi) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-Avi, C-His labeled tag.
LILRB1/CD85j/ILT2 Protein is an inhibitory receptor broadly expressed on leukocytes and recognizes HLA-class I and human cytomegalovirus UL18. LILRB1/CD85j/ILT2 Protein, Rhesus macaque (HEK293, Fc) is the recombinant Rhesus Macaque-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-hFc labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Cynomolgus (HEK293, His) is the recombinant cynomolgus-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-His labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Domain1&2 Protein, Human (HEK293, His) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-His labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Human (346a.a, HEK293, His) is the recombinant human-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-His labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Rhesus Macaque (474a.a, HEK293, Fc) is the recombinant Rhesus Macaque-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-hFc labeled tag.
The LILRB1/CD85j/ILT2 protein is a receptor for class I MHC antigens and recognizes multiple HLA alleles and H301/UL18 (human cytomegalovirus MHC homolog). Ligand binding induces inhibitory signals that downregulate immune responses. LILRB1/CD85j/ILT2 Protein, Rhesus macaque (474a.a, HEK293, His) is the recombinant Rhesus Macaque-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-His labeled tag.
LILRB1/CD85j/ILT2 Protein is an inhibitory receptor broadly expressed on leukocytes and recognizes HLA-class I and human cytomegalovirus UL18. LILRB1/CD85j/ILT2 Protein, Rhesus Macaque (HEK293, His) is the recombinant Rhesus Macaque-derived LILRB1/CD85j/ILT2 protein, expressed by HEK293 , with C-6*His labeled tag.
hsa-miR-7-5p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
hsa-miR-7-5p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-2-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7a-5p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
hsa-miR-7-1-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7b-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7a-2-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
mmu-miR-7b-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
mmu-miR-7a-2-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-5p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
rno-miR-7b antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-5p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-5p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-5p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
LILRB1 Human Pre-designed siRNA Set A contains three designed siRNAs for LILRB1 gene (Human), as well as a negative control, a positive control, and a FAM-labeled negative control.
rno-miR-7b mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
rno-miR-7b inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
rno-miR-7a-1-3p mimics are small, chemically synthesized double-stranded RNAs that mimic endogenous miRNAs and enable miRNA functional analysis by up-regulation of miRNA activity.
rno-miR-7b agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-5p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-2-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-1-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
rno-miR-7a-1-3p inhibitors are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA inhibitors have full-length nucleotide 2'-methoxy modification. The miRNA inhibitors strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning.
hsa-miR-7-1-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-2-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7b-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-2-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
rno-miR-7a-1-3p agomirs are chemically-modified double-strand miRNA mimics with modified mature miRNA strand: 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 3' end cholesterol group, and full-length nucleotide 2'-methoxy modification. They are designed to mimic endogenous miRNAs and recommended for miRNA functional studies. Compared with miRNA mimics, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-1-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7b-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
hsa-miR-7-2-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
mmu-miR-7a-2-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
rno-miR-7a-1-3p antagomirs are chemically-modified oligonucleotides that hybridize with mature miRNAs. The miRNA antagomirs have 2 phosphorothioates at the 5' end, 4 phosphorothioates at the 3' end, 1 cholesterol group at the 3' end, and full-length nucleotide 2'-methoxy modification. The miRNA antagomirs strongly compete with mature miRNAs to prevent the complementary pairing of miRNAs and their target genes, thereby inhibiting miRNAs from functioning. Stability of miRNA antagomirs appears to be significantly higher than miRNA inhibitors, they exhibits enhanced cellular uptake, stability and regulatory activity in vivo.
Inquiry Online
Your information is safe with us. * Required Fields.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
MedchemExpress Validation 03
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
MedchemExpress Validation 04
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedChemExpress values your privacy and your trust is important to us. We use cookies to enhance your website experience. Some cookies are necessary to run the website.
Privacy and Cookie Policy