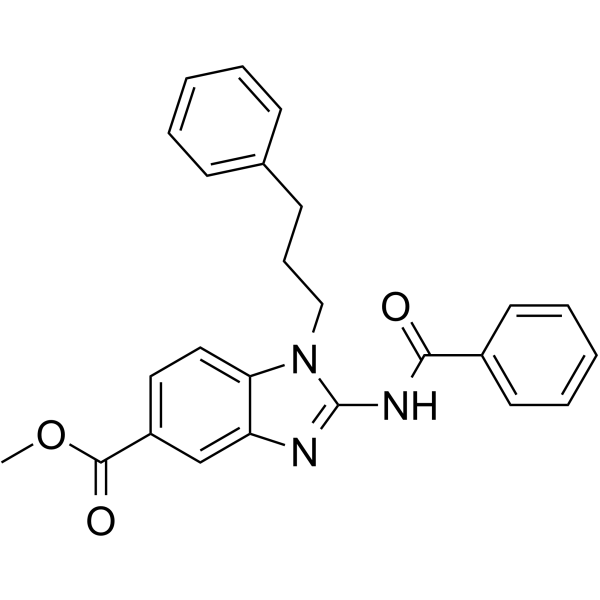
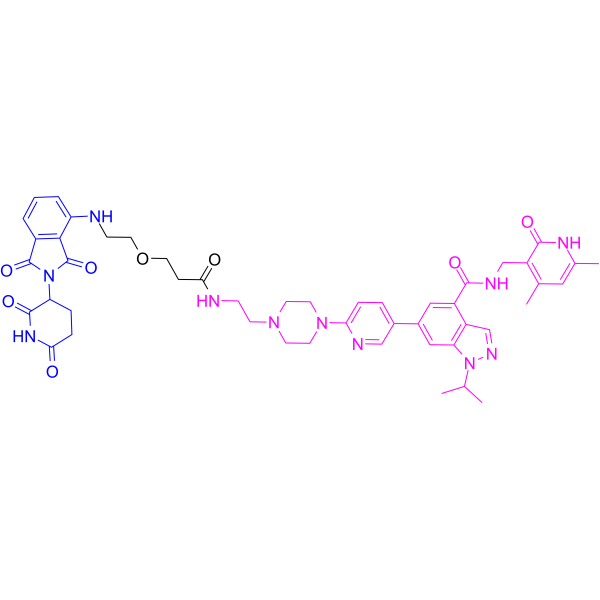
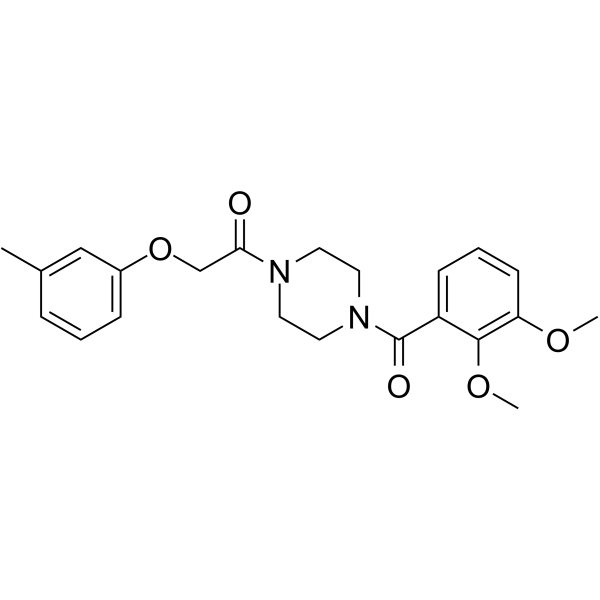
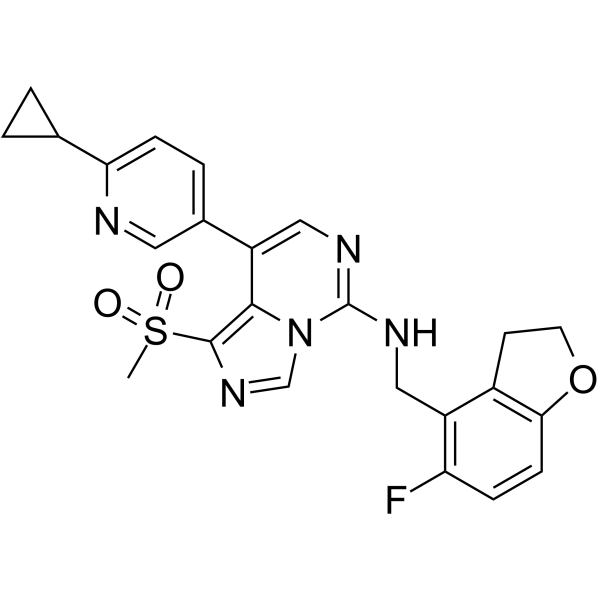
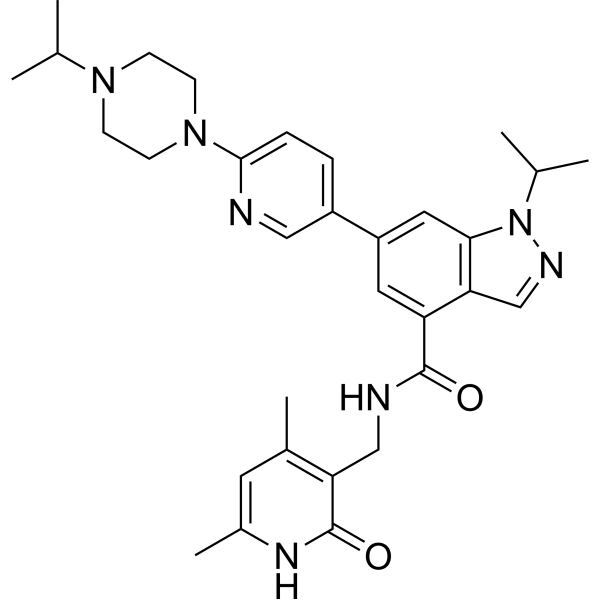
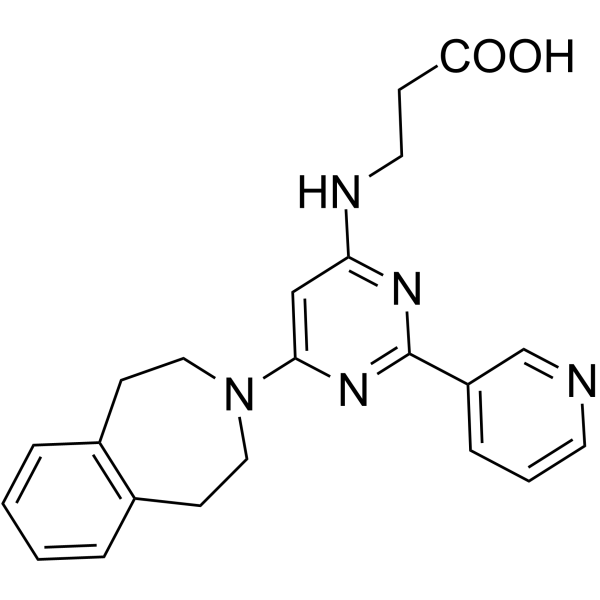
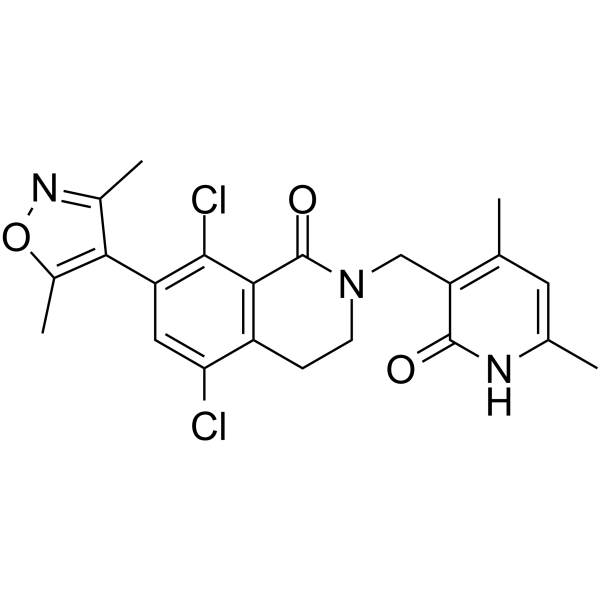
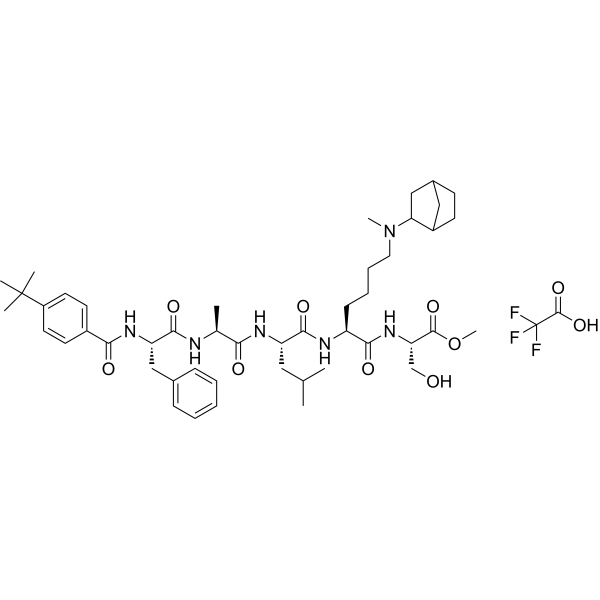
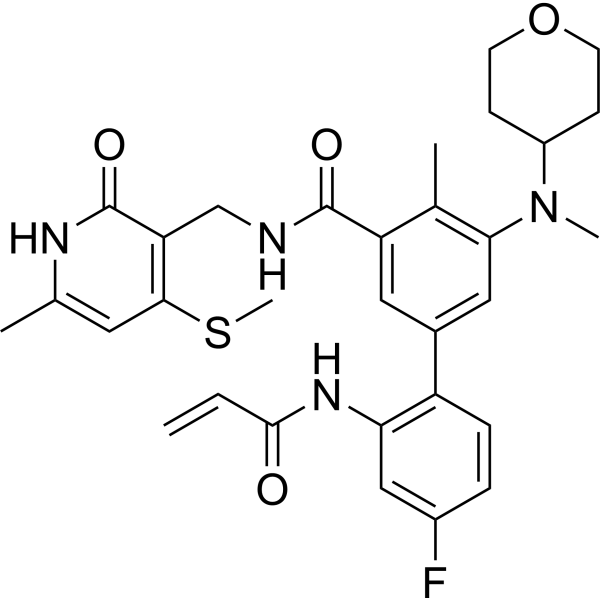
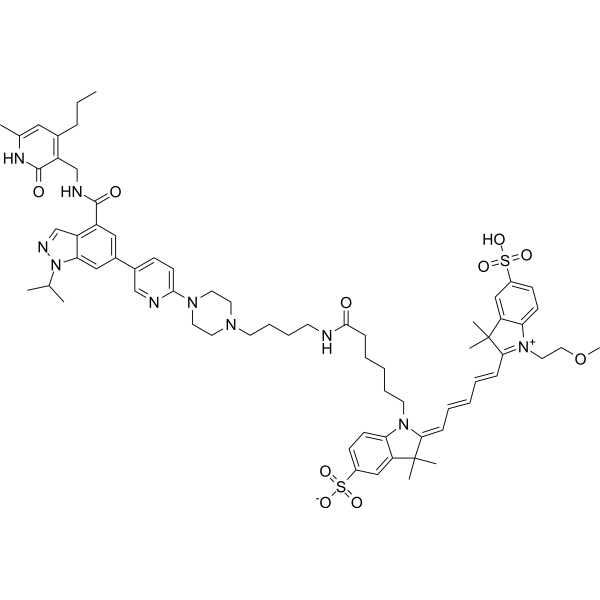
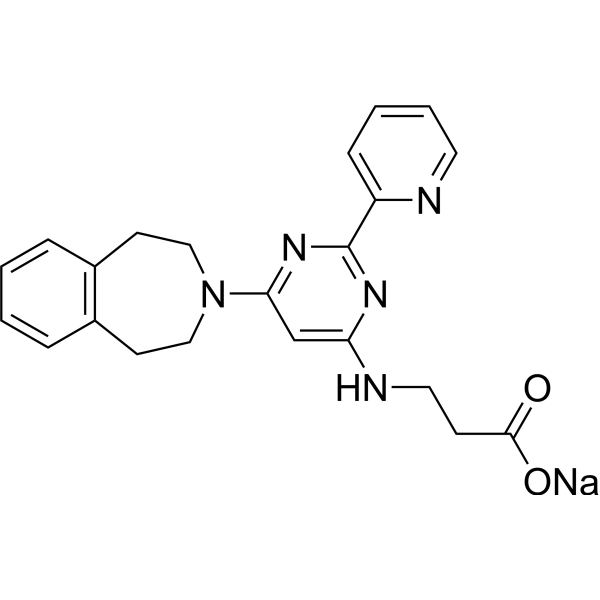
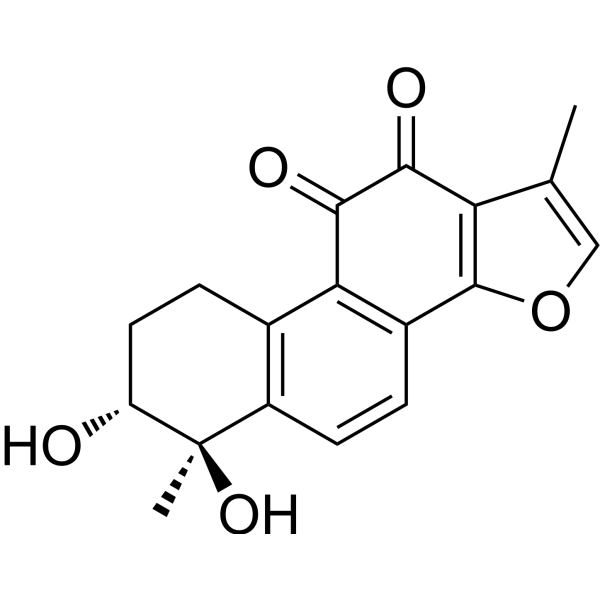
From 11:00 pm to 12:00 pm EST ( 8:00 pm to 9:00 pm PST ) on January 6th, the website will be under maintenance. We are sorry for the inconvenience. Please arrange your schedule properly.
GSK-J4 is a potent dual inhibitor of H3K27me3/me2-demethylasesJMJD3/KDM6B and UTX/KDM6A with IC50s of 8.6 and 6.6 μM, respectively. GSK-J4 inhibits LPS-induced TNF-α production in human primary macrophages with an IC50 of 9 μM. GSK J4 is a cell permeable proagent of GSK-J1 . GSK-J4 induces endoplasmic reticulum stress-related apoptosis .
GSK-J4 hydrochloride is a potent dual inhibitor of H3K27me3/me2-demethylasesJMJD3/KDM6B and UTX/KDM6A with IC50s of 8.6 and 6.6 μM, respectively. GSK-J4 hydrochloride inhibits LPS-induced TNF-α production in human primary macrophages with an IC50 of 9 μM. GSK-J4 hydrochloride is a cell permeable proagent of GSK-J1 .
BRD4770 is a histone methyltransferase G9a inhibitor. BRD4770 reduces di- and trimethylation of lysine 9 on histone H3 (H3K9) with an EC50 of 5 μM, and has less or little effect toward H3K27me3, H3K36me3, H3K4me3, and H3K79me3. BRD4770 can activate the ataxia telangiectasia mutated (ATM) pathway and induce cell senescence .
MS177 is an effective and fast-acting EZH2 degrader. MS177 is a PROTAC that consists of a CRBN ligand, linker, and a potent enzymatic EZH2 inhibitor C24 (C24 IC50): 12 nM). MS177 effectively depletes both canonical EZH2-PRC2 and noncanonical EZH2-cMyc complexes. MS177 induces leukaemia cell growth inhibition, apoptosis and cell cycle progression arrest .
MS37452 is a potent inhibitor of CBX7 chromodomain binding to H3K27me3, with a Kd of 27.7 μM. MS37452 can derepress transcription of polycomb repressive complex target gene p16/CDKN2A by displacing CBX7 binding to the INK4A/ARF locus in prostate cancer cells .
EEDi-5285 is an exceptionally potent and orally active embryonic ectoderm development (EED) inhibitor with an IC50 value of 0.2 nM for binds to the EED protein. EEDi-5285 has anti-cancer activity .
EZH2-IN-14 is a selective EZH2 (Histone Methyltransferase) inhibitor with an IC50 of 12 nM. EZH2-IN-14 inhibits the methyltransferase activity of EZH2/PRC2 (that is, reducing H3K27me3). EZH2-IN-14 shows >200-fold selective for EZH2 over the highly homologous H3K27 methyltransferase EZH1 .
GSK-J2 is an isomer of GSK-J1 that does not have any specific activity. GSK-J1 is a potent inhibitor of H3K27me3/me2-demethylases JMJD3/KDM6B and UTX/KDM6A.
H3K27(Me3) (15-34), a histone peptide, is a repressive chromatin mark derived from human histone. Polycomb Repressive Complex 2 (PRC2) is a multiprotein complex that catalyzes the methylation of H3K27(Me) .
PF-06726304 is a potent and selective EZH2 inhibitor. PF-06726304 inhibits wild-type and Y641N mutant EZH2 with Kis of 0.7 and 3.0 nM, respectively. PF-06726304 displays robust antitumor growth activity .
UNC4976 TFA is a positive allosteric modulator (PAM) peptidomimetic of CBX7 chromodomain binding to nucleic acids. UNC4976 TFA simultaneously antagonizes H3K27me3-specific recruitment of CBX7 to target genes while increasing non-specific binding to DNA and RNA .
SPEN-IN-1 (compound X1) is an inhibitor of SPEN which is a protein factor with a Kd value of 47 nM. SPEN-IN-1 has high selectivity for RepA, a 431-nucleotide domain in Xist (a non-coding RNA prototype) that comprises 8.5 units of a GC-rich motif responsible for gene silencing .
JQEZ5 is a potent and selective EZH2 lysine methyltransferase inhibitor. JQEZ5 SAM-competitively inhibits polycomb repressive complex 2 (PRC2) with an IC50 of 80 nM. JQEZ5 has anti-tumor effects .
UNC4976 is a positive allosteric modulator (PAM) peptidomimetic of CBX7 chromodomain binding to nucleic acids. UNC4976 simultaneously antagonizes H3K27me3-specific recruitment of CBX7 to target genes while increasing non-specific binding to DNA and RNA .
NPD13668 is an EZH2-mediated gene silencing modulator. NPD13668 inhibits EZH2/PRC2 (polycomb repressive complex 2) activity. NPD13668 reactivates the expression of silenced H3K27me3 target genes together with depletion of the H3K27me3 modification. NPD13668 can be used for the study of prostate cancer and ovarian cancer .
WQQ-345 is an orally active BCAT1 inhibitor with an IC50 values of 10.8 mM. WQQ-345 reduces cellular α-KG levels, upregulating H3K27me3 expression, decreasing glycolytic enzyme expression, and impairing glycolysis activity. WQQ-345 reduces colony formation, suppresses growth of BCAT1-high TKI-resistant lung cancer cells. WQQ-345 exerts in vitro and in vivo antitumor activity. WQQ-345 can be used for the research of TKI-resistant non-small cell lung cancer and TKI-resistant lung cancer .
PROTAC EZH2 Degrader-3 (compound ZJ-20) is a EZH2 PROTAC degrader. PROTAC EZH2 Degrader-3 (5 μM, 24 h) not only potently inhibits the expression of EZH2 protein, but also had a strong inhibits effect on the expression levels of other subunits of PRC2 as well as H3k27me3 protein. PROTAC EZH2 Degrader-3 shows anti-proliferative activity and blocks the cell cycle in the G0-G1 phase and induces cell apoptosis((Blue: cereblon ligand Pomalidomide (HY-10984), Black: linker HY-W361751;Pink: EZH2 inhibitor Tazemetostat (HY-13803)) .
IHMT-EZH2-426 (compound 38) is an effective covalent EZH2 degrader, with IC50 values of 1.3 nM, 1.2 nM, and 1.7-3.5 nM for EZH2 wild type, EZH2-A687V, and EZH2-Y641F/Y641N/Y641S, respectively. IHMT-EZH2-426 reduces H3K27me3 and EZH2 levels and shows effective anti-proliferative effects in B-cell lymphoma and TNBC cell lines.
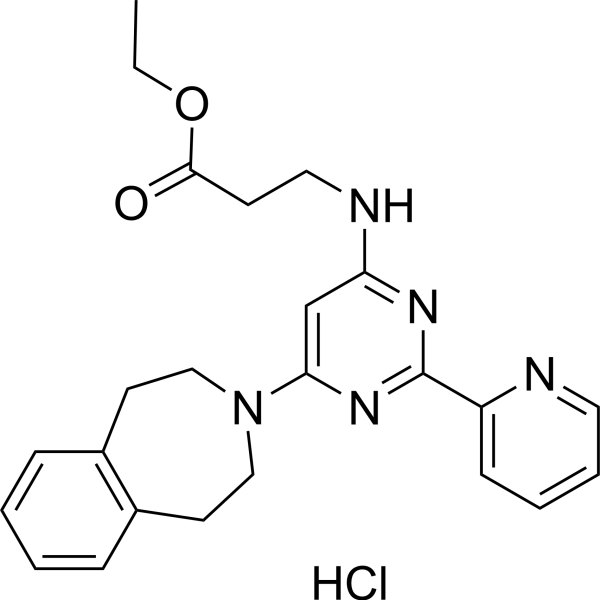
GSK-J4 (hydrochloride) (Standard) is the analytical standard of GSK-J4 (hydrochloride). This product is intended for research and analytical applications. GSK-J4 hydrochloride is a potent dual inhibitor of H3K27me3/me2-demethylases JMJD3/KDM6B and UTX/KDM6A with IC50s of 8.6 and 6.6 μM, respectively. GSK-J4 hydrochloride inhibits LPS-induced TNF-α production in human primary macrophages with an IC50 of 9 μM. GSK-J4 hydrochloride is a cell permeable proagent of GSK-J1 .
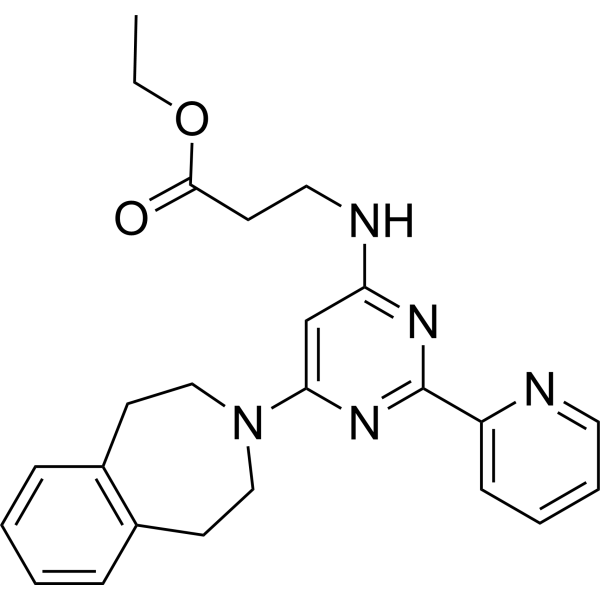
GSK-J4 (Standard) is the analytical standard of GSK-J4. This product is intended for research and analytical applications. GSK-J4 is a potent dual inhibitor of H3K27me3/me2-demethylases JMJD3/KDM6B and UTX/KDM6A with IC50s of 8.6 and 6.6 μM, respectively. GSK-J4 inhibits LPS-induced TNF-α production in human primary macrophages with an IC50 of 9 μM. GSK J4 is a cell permeable proagent of GSK-J1 . GSK-J4 induces endoplasmic reticulum stress-related apoptosis .
DC-PRC2in-01 is a potent EZH2-EED interaction inhibitor with an IC50 of 4.21 μM and a Kd of 4.56 μM. DC-PRC2in-01 disrupts the EZH2-EED interaction, leading to degradation of PRC2 core proteins and decrease of H3K27me3 levels, inhibition of PRC2-driven lymphoma cell proliferation, and cell cycle arrest. DC-PRC2in-01 can be used for the research of PRC2-related cancers, such as Diffuse Large B-cell Lymphoma (DLBCL) and follicular lymphoma (FL) .
Tanshindiol C is a S-adenosylmethionine-competitive EZH2 (Histone Methyltransferase) inhibitor with an IC50 of 0.55 μM for inhibiting the methyltransferase activity. Tanshindiol C is also an activator of both Nrf2 and Sirtuin 1 (Sirt1) in macrophages. Tanshindiol C possesses anti-cancer activity, and can be used for atherosclerosis research .
GSK-J2 sodium is the sodium form of GSK-J2 (HY-15648A). GSK-J2 is an isomer of GSK-J1, and does not have any specific activity. GSK-J1 (HY-15648) is a potent inhibitor of H3K27me3/me2-demethylases JMJD3/KDM6B and UTX/KDM6A .
EZH2-IN-1 (compound 3) is a selective and SAM-competitive EZH2 and EZH1 inhibitor with an IC50s of 32 nM, 197 nM and 213 nM for EZH2wt, EZH2 Y641N mutant and EZH1, respectively. EZH2-IN-1 reduces bulk H3K27me3 and H3K27me2 levels. EZH2-IN-1 has the potential for diffuse large B cell lymphoma research .
EED-IN-3 is an orally active EED inhibitor. EED-IN-3 effectively inhibits PRC2 by binding to EED (IC50 = 0.62 μM for EED) and downregulates H3K27me3. EED-IN-3 can efficiently and selectively inhibit PC3 cells with the IC50 of 3.69 μΜ and could significantly suppress colony formation and migration. EED-IN-3 can be used for research on prostate cancer.
D-01 is a dual-targeting inhibitor of HIF-1α and EZH2 (IC50: 4.86 μM and 0.99 μM respectively). D-01 inhibits the expression of H3K27me3 protein. D-01 inhibits the migration, clone and the invasion of A549 cells, and also inhibits tube formation of HUVECs. D-01 can be used for research of lung cancer .
HDM-IN-1 (Compound A4) is an inhibitor for fungal histone demethylase (HDM), that inhibits the H3K27me3 in Cryptococcus neoformans and in Candida auris with IC50s of 134 and 12 nM. HDM-IN-1 exhibits inhibitory efficacy against C. neoformans and C. auris with MIC80 of 0.5-2 μg/mL, through inhibition of the biofilm and capsule formation. HDM-IN-1 exhibits antifungal activity in ICR mouse model .
TDI-11904 is a peripherally active EZH2 inhibitor with an EZH2IC50 of 0.9 nM. TDI-11904 inhibits EZH2 methyltransferase activity, reduces the trimethylation level of H3K27me3, and decreases intracellular H3K27me3 content. TDI-11904 inhibits the proliferation of lymphoma cells. TDI-11904 can be used for the research of lymphoma .
EZH2-IN-7 is a potent inhibitor of EZH2. EZH2 overexpression or mutations in the SET region (Y641F, Y641N, A687V, A677G point mutations) all lead to abnormal elevation of H3K27me3 and promote the growth and development of many types of tumors, such as breast cancer, prostate cancer, leukemia, etc. EZH2-IN-7 has the potential for the research of cancer diseases (extracted from patent WO2021129629A1, compound 259) .
CBX2-IN-1 is a CBX2 inhibitor with an IC50 of 9.6 μM. CBX2-IN-1 binds the CBX2 chromodomain’s H3K27me3 binding channel, blocks interaction between CBX2 and H3K27me3, and exhibits selective affinity toward CBX2, CBX4, CBX6, CBX7, and CBX8 over CBX1, CBX3, and CBX5. CBX2-IN-1 can be used for the research of prostate cancers .
H3K27(Me) (15-34), a histone peptide, is a repressive chromatin markderived from human histone. Polycomb Repressive Complex 2 (PRC2) is a multiprotein complex that catalyzes the methylation of H3K27(Me) .
EED-IN-5 is an orally active, EZH2-selective trisubstituted pyridine-based EED-H3K27me3 inhibitor and immunomodulator with anti-inflammatory activity. The IC50 value of EED-IN-5 against EED is 28.21 nM. In mouse models, EED-IN-5 preferentially and persistently accumulates in lymph nodes after oral administration. By reducing the H3K27me3 level of dendritic cells and inhibiting their migration, EED-IN-5 decreases the infiltration of specific dendritic cells, macrophages and T cells into the spinal cord and brain. EED-IN-5 exhibits hERG inhibitory activity, shows negative results in the Mini-Ames test, and causes no obvious toxicity upon long-term high-dose administration. EED-IN-5 can be used for the research of multiple sclerosis .
EED-IN-4 is an orally active, EZH2-selective immunomodulator and EED-H3K27me3 inhibitor (EED, IC50=28.21 nM) with anti-inflammatory activity. In mouse models, EED-IN-4 preferentially and persistently accumulates in lymph nodes after oral administration. By reducing the H3K27me3 level of dendritic cells and inhibiting their migration, EED-IN-4 reduces the infiltration of immune cells into the central nervous system and effectively alleviates spinal cord inflammation. EED-IN-4 shows weak inhibitory activity against hERG channels and is non-mutagenic, with no obvious toxicity observed upon long-term oral administration. EED-IN-4 can be used for the research of multiple sclerosis .
H3K27(Me2) (15-34), a histone peptide, is a repressive chromatin mark derived from human histone. Polycomb Repressive Complex 2 (PRC2) is a multiprotein complex that catalyzes the methylation of H3K27(Me) .
PROTAC EZH2 Degrader-44 (compound 60) is a highly efficient PROTAC degrader targeting the EZH2-PRC2 complex. By recruiting the CRBN E3 ligase and relying on the proteasome system, PROTAC EZH2 Degrader-44 simultaneously induces the degradation of core components EZH2, SUZ12 and EED, thereby significantly reducing the levels of H3K27me3 and CARM1. PROTAC EZH2 Degrader-44 exerts antiproliferative effects through a dual mechanism: on the one hand, it triggers mitochondrial dysfunction leading to decreased membrane potential; on the other hand, it strongly promotes apoptosis by regulating Bcl-2 family proteins (upregulating Bax, Caspase-3 and PARP, and downregulating Bcl-2). PROTAC EZH2 Degrader-44 exhibits only extremely low cytotoxicity in human normal mammary epithelial, liver and kidney cells, showing a favorable safety window. PROTAC EZH2 Degrader-44 is an ideal tool molecule for exploring the mechanisms of targeted therapy for triple-negative breast cancer .
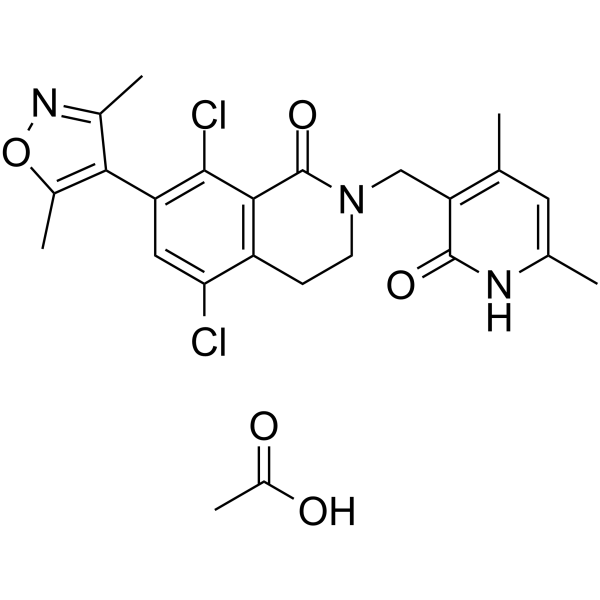
PF-06726304 acetate is a potent and selective EZH2 inhibitor. PF-06726304 acetate inhibits wild-type and Y641N mutant EZH2 with Kis of 0.7 and 3.0 nM, respectively. PF-06726304 acetate displays robust antitumor growth activity .
SKLB-03220 is a selective and orally active EZH2 covalent inhibitor with an IC50 of 1.72 nM for EZH2 MUT. SKLB-03220 exhibits weak activities against other tested histone methyltransferases (HMTs) and kinases. SKLB-03220 displays noteworthy potency against ovarian cancer cell lines and induces cell apoptosis. SKLB-03220 significantly inhibits tumor growth in PA-1 xenograft model. SKLB-03220 can be used for the study of ovarian cancer .
PROTAC EZH2 Degrader-9 is orally active EZH2 PROTAC degrader degrading EZH2 via the ubiquitin-proteasome pathway. PROTAC EZH2 Degrader-9 downregulates PRC2 core subunits and potent inhibition of H3K27me3 without affecting common CRBN neosubstrates while it was selective over GSp'T1 and ikZF1/3. PROTAC EZH2 Degrader-9 exhibits potent antiproliferative activity against multiple cancer cell lines by inducing cell cycle and apoptosis. PROTAC EZH2 Degrader-9 reverses PRC2-mediated gene silencing and inhibiting EZH2 non-catalytic target gene activation. PROTAC EZH2 Degrader-9 can be used for leukemia, lymphoma, and non-small cell lung cancer research .
EZH2-IN-4 is an orally active, potent EZH2 inhibitor with IC50s of 0.923 nM and 2.65 nM against wild type (WT) 5-membered (5-mer) EZH2 and mutant 5-mer EZH2, respectively. EZH2-IN-4 has anti-cancer activity .
JQEZ5 (Standard) is the analytical standard of JQEZ5 (HY-100846). This product is intended for research and analytical applications. JQEZ5 is a potent and selective EZH2 lysine methyltransferase inhibitor. JQEZ5 SAM-competitively inhibits polycomb repressive complex 2 (PRC2) with an IC50 of 80 nM. JQEZ5 has anti-tumor effects .
PROTAC EZH2 Degrader-34 (compound E7 (13)) is an EZH2-target PROTAC degrader with a human EZH2IC50 of 6.30 μM. PROTAC EZH2 Degrader-34 can be used for the research of pfeiffer cancer, prostate cancer .
H3K27(Me3) (15-34), a histone peptide, is a repressive chromatin mark derived from human histone. Polycomb Repressive Complex 2 (PRC2) is a multiprotein complex that catalyzes the methylation of H3K27(Me) .
UNC4976 TFA is a positive allosteric modulator (PAM) peptidomimetic of CBX7 chromodomain binding to nucleic acids. UNC4976 TFA simultaneously antagonizes H3K27me3-specific recruitment of CBX7 to target genes while increasing non-specific binding to DNA and RNA .
UNC4976 is a positive allosteric modulator (PAM) peptidomimetic of CBX7 chromodomain binding to nucleic acids. UNC4976 simultaneously antagonizes H3K27me3-specific recruitment of CBX7 to target genes while increasing non-specific binding to DNA and RNA .
H3K27(Me) (15-34), a histone peptide, is a repressive chromatin markderived from human histone. Polycomb Repressive Complex 2 (PRC2) is a multiprotein complex that catalyzes the methylation of H3K27(Me) .
H3K27(Me2) (15-34), a histone peptide, is a repressive chromatin mark derived from human histone. Polycomb Repressive Complex 2 (PRC2) is a multiprotein complex that catalyzes the methylation of H3K27(Me) .
Tanshindiol C is a S-adenosylmethionine-competitive EZH2 (Histone Methyltransferase) inhibitor with an IC50 of 0.55 μM for inhibiting the methyltransferase activity. Tanshindiol C is also an activator of both Nrf2 and Sirtuin 1 (Sirt1) in macrophages. Tanshindiol C possesses anti-cancer activity, and can be used for atherosclerosis research .
TriMethyl-Histone H3 (Lys27) Antibody (YA030) is a Rabbit-derived and non-conjugated IgG monoclonal antibody, targeting to TriMethyl-Histone H3 (Lys27).
TriMethyl-Histone H3 (Lys27) Antibody (YA9825) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to TriMethyl-Histone H3 (Lys27).
WB, ICC/IF, IHC-P, IF-Tissue, FC, ChIP, Dot Blot, IP
Human, Mouse, Rat
Acetyl-Histone H3 (Lys9) Antibody (YA9827) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Acetyl-Histone H3 (Lys9).
DiMethyl-Histone H3 (Lys27) Antibody (YA9831) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to DiMethyl-Histone H3 (Lys27).
MonoMethyl-Histone H3 (Lys4) Antibody (YA9833) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to MonoMethyl-Histone H3 (Lys4).
MonoMethyl-Histone H3 (Lys79) Antibody (YA9839) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to MonoMethyl-Histone H3 (Lys79).
TriMethyl-Histone H3 (Lys36) Antibody (YA9840) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to TriMethyl-Histone H3 (Lys36).
MonoMethyl-Histone H3 (Lys9) Antibody (YA9826) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to MonoMethyl-Histone H3 (Lys9).
Phospho-Histone H3 (Ser28) Antibody (YA9829) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Phospho-Histone H3 (Ser28).
Acetyl-Histone H3 (Lys4) Antibody (YA9830) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Acetyl-Histone H3 (Lys4).
Acetyl-Histone H3 (Lys23) Antibody (YA9832) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Histone H3 (acetyl Lys23).
Acetyl-Histone H3 (Lys36) Antibody (YA9834) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Acetyl-Histone H3 (Lys36).
Phospho-Histone H3 (Ser10) Antibody (YA9835) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Phospho-Histone H3 (Ser10).
DiMethyl-Histone H3 (Lys4) Antibody (YA9836) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to DiMethyl-Histone H3 (Lys4).
MonoMethyl-Histone H3 (Lys14) Antibody (YA9837) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to MonoMethyl-Histone H3 (Lys14).
Mono+Di+TriMethyl-Histone H3 (Lys36) Antibody (YA9828) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Mono+Di+TriMethyl-Histone H3 (Lys36).
Mono+Di+TriMethyl-Histone H3 (Lys4) Antibody (YA9838) is a Rabbit-derived and non-conjugated IgG Recombinant, Monoclonal antibody, targeting to Mono+Di+TriMethyl-Histone H3 (Lys4).
TriMethyl-Histone H3 (Lys27) Antibody (YA030) is a Rabbit-derived and non-conjugated IgG monoclonal antibody, targeting to TriMethyl-Histone H3 (Lys27).
(5)
Product Comparison
Compare
Clear All
Compare Products
Products
In-stock
-
+
Add to Cart
Cat. No.
Host
Reactivity
Application
Dilution Ratio
Molecular Weight
Conjugation
Clonality
Immunogen
Appearance
Isotype
Gene ID
SwissProt ID
Purity
Formulation
Free Sample
YesNo
Size
* This product has been "discontinued".
Optimized version of product available:
/
In-stock
-
+
Add to Cart
Get quote
Inquiry Online
Your information is safe with us. * Required Fields.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
MedchemExpress Validation 03
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
MedchemExpress Validation 04
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedchemExpress Validation
Western blot analysis of extracts from THP-1(lane 2(20μg), Jurkat (lane 3(20μg) and NIH3T3(lane 4(20μg) using FOXO1A (HY-P80132) Rabbit mAb. Proteins were transferred
to a PVDF membrane and blocked with 5% non-fat milk in TBST for 2 hour at room temperature. The primary antibody (1/1000) and Loading control antibody (Beta Actin, HY-P80438, 1/10000) was
used in 5% non-fat milk in TBST at 4°C overnight. Goat Anti-Mouse/Rabbit IgG-HRP Secondary Antibody (1/10000) was used for 1 hour at room temperature.
MedChemExpress values your privacy and your trust is important to us. We use cookies to enhance your website experience. Some cookies are necessary to run the website.
Privacy and Cookie Policy